 设为首页
加入收藏
设为首页
加入收藏
扩展资料
生物拟态
拟态是指某些生物在进化过程中形成的外表形状或色泽斑与其它生物或非生物异常相似的现象,是生物适应环境的最为典型的例子。随着生态学研究的不断深入,人们发现动物甚至植物拟态的现象是十分普遍的,拟态在许多生态系统中起着十分重要的作用。
1 拟态的发现
一种生物能够模拟另一种生物以便欺骗敌人,逃避敌害,这种观点一个多世纪以来一直吸引着生物学家们。这种特殊的生存策略是由英国博物学家Henrry W. Bates在1862年首先描述的。他观察到南美蝴蝶中某些具鲜艳色彩的种类从外形上看几乎完全相同,但有些是有毒的,而另外一些却是完全无毒的,他由此推测这些无毒的生物伪装成有毒的,以便逃避被捕食的厄运。他称这种现象为拟态,这就是著名的贝氏拟态。动物的拟态相当普遍,人们研究得也较多;而植物拟态远不如动物拟态普遍,因此很长一段时间未被人们注意。事实上,早在1793年德国博物学家 Christian K. Sprengel就观察到植物象动物一样,可以模拟其它生物。他发现某些植物特别是兰科植物的某些种类自己不能分泌花蜜,但它们能模拟生活在同一环境中的可以产生花蜜的植物的形态。直到最近,随着植物学家对植物与其它生物间关系研究的深入,特别是对热带生态系统研究,人们才认识到植物拟态比预想的要广泛得多。
2 拟态的组成
不论是动物中的拟态还是植物中的拟态都是一个由三部分组成的系统。一是模型,是被模拟动物、植物或其它生物或非生物;二是拟态生物;三是信号接受者或被欺骗者。拟态生物模拟的内容包括形态结构、颜色花纹、行为方式或其它使拟态生物与模型相似的属性。这里模型可以是与拟态生物无任何联系的其它生物或非生物,如某生物长期生活背景的衬托物等。枯叶蝶与枯叶异常相似,在这一拟态系统中,枯叶为模型,枯叶蝶为拟态生物,某些食虫鸟则为信号接受者或被欺骗者。值得一提的是,信号接受者之所以会被欺骗,与其眼睛及接受拟态生物所反射来的光有密切关系。例如瑞典乌普沙拉系统植物学会的L. Anders Nilsson发现,两种独居蜂 Chelostoma fuliginosum 和 C. campanutarum 同时光顾多种钟形花(bellflouer,又叫风铃草,风铃草属)和兰科植物 Cephalanthera rubra(头蕊兰属)。钟形花与兰花形态和颜色(兰花是玫瑰红色,钟形花是兰紫色的)都不一样,更重要的是钟形花为来访者提供了丰富的花蜜,而兰花根本不产生花蜜。为什么看上去与钟形花不同也不分泌花蜜的兰花却能吸引独居蜂为其传粉呢?Nilsson利用光谱仪解决了这个问题,他发现虽然在人眼里兰花与钟形花颜色不同,但两种花反射的光曲线轮廊在独居蜂视觉范围内却是完全相同的,它们对紫外线末端最敏感,象许多昆虫一样,它们不能区别多种不同的红色。因此对独居蜂来说,兰花和钟形花是完全相同的,兰花在此实际上是拟态生物,钟形花充当了模型的角色,独居蜂则是被欺骗者。
3 拟态的类型
生物的拟态可以分为两种类型,一是贝氏拟态。即由模型、拟态生物和信号接受者三部分组成的系统。例如尺蠖的幼虫在树皮上斜伸不动,和小树枝一样。这种拟态的特点是一种生物由于模拟另一种生物而获得了生存优势;另外一种拟态,是著名的缪勒拟态是19世纪德国动物学家Fritz M{ller命名的,他首先描述了这种现象。在这种拟态类型中,许多没有联系的生物在形态和行为上彼此相似,形成看上去是一群的效果,从而得到好处。由于在这一拟态系统中没有欺骗,模型与拟态生物没有明显区别,无所谓谁模拟谁的问题,所有参与者贡献相同,共同获益,这些与贝氏拟态显然不同。因此许多生物学家认为这不是真正的拟态,只是生活在相同环境中的生物以相似的方式进化的结果(即趋同进化,典型的例子是沙漠中肉质植物的进化)。
朱晓林,生物学通报,1995年第1期
扩展资料
保护色、警戒色、拟态的区别
教学实践发现:学生解答下面的问题常会出错。“选出下列不属于拟态的项:A.枯叶蝶似枯叶。B.有的无毒蛇具有毒蛇的鲜艳体色。C.南美蚕蛾幼虫的体态、色斑似眼镜蛇。D.美洲棕王蝶幼虫有毒,体节有棕色大斑纹。”答案应选D,但有的学生却不知B、C两项为何不能选。而下面一题学生解答起来更是疑惑不定。“下列分别属于保护色和警戒色的一组是:A.虎具有斑纹和黄蜂腹部的条纹。B.昆虫的体色和北极熊的体色。C.枯叶蝶具有的特征和黄刺蛾幼虫的鲜艳色彩。D.腹蛇体表的斑纹和比目鱼的体色。”答案应选A,而不少学生判断出选A后却感到疑惑不定,有的甚至改选C。
为什么会出现上述情况呢?主要是学生没有准确地把握保护色、警戒色、拟态这三个概念的含义,不能正确区分三种现象,特别是对拟态和保护色混淆不清。
如何才能有效区别这三种现象呢?还得从概念的含义出发。
保护色和拟态现象都表现为与环境色彩相似,不易被识别,而警戒色则表现得与环境不同,容易被发现,且具警戒色的动物一般都具有潜在的伤害性,据此可把警戒色与其他两者区分开。保护色表现为与环境色彩相似,这里的“环境色彩”应是环境中主要的占优势的色彩,如春夏的草坪是绿色,冬天的雪地是白色;拟态是与环境中某种生物或非生物相似,而这种生物或非生物的颜色等特征并不一定在环境中占优势,并非主要色彩。而且拟态现象往往与生物所处的运动状态有关,一般是处于相对静止状态时才相似,一旦运动起来则就不相似了;而保护色则与运动状态基本无关,如枯叶蝶停息在树枝上的模样像枯叶,“停息”状态才像枯叶,一旦飞舞起来就不像了而我们捕捉昆虫也许都有这种体验:有时看到昆虫由这里飞向另一个地方,但马上在另一个地方搜寻,却不能立即找到。
基于以上分析,区别这三种现象就容易多了。如上面提到的“有的无毒蛇具有毒蛇的鲜艳体色”、“南美蚕蛾幼虫的体态、色斑似眼镜蛇”,这两者都是与环境中某种生物相似,故应是拟态现象,而“虎具有的斑纹”是与环境色彩相似,应是保护色。再如下面几种现象:“A.避役的体色能够随环境色彩的变化而改变并与环境的色彩保持一致。B.生活在亚马逊河流域的南美鲈鱼形如败叶,浮在水面。C.黄蜂腹部具有黄黑相间的条纹。D.腹蛇体表的斑纹。E.生活在海藻中的一种鱼,遇到敌害时,身体侧立不动,似一株植物,从而逃避敌害。”这里的黄蜂、腹蛇具潜在伤害性,且条纹、斑纹很醒目,故C、D是警戒色;避役始终与环境色彩一致,不易被发现,故A是保护色;南美鲈鱼和海藻中的那种鱼都是与环境中某种生物相似,而且只有在“侧立不动”或“浮在水面”不动时才似植物、如败叶,一旦游动起来肯定就不像了,故B、E是拟态。
覃鸿举,生物学教学,1999年第12期
扩展资料
生物的竞争与适应
1 物种竞争
从达尔文时代的“生存竞争,优胜劣汰”的理论产生到现在,生态学家在关于生物竞争方面进行了广泛而深入的研究。所谓“竞争”,是指种内或种间的两个或更多个个体间,由于它们的需求或多或少地超过了共同资源的供应而产生的一种生存斗争现象,所以在这些竞争着的个体间相互施加着不利的影响。这种影响可以表现为存活率、个体增加速率、体重和生育力的下降,而其总的效应则表现为种群增殖率随密度增长而下降。当简单生活史的单种种群在无限环境中时,其种群数量应呈指数式增长。然而,我们所拥有的地球可供生物的资源是有限的,因此,各种生物为了自身的生存,必然会与其他个体对资源进行竞争。Verhulst (1838)和Pearl与Reed(1920)最早彼此独立地用数学式表达物种在有限环境中种群所呈现的逻辑斯谛增长形式,该模型表明,随着种群内个体数量的增多,对有限资源的种内竞争也将加剧,从而使种群不能充分实现其内禀增长能力所允许的增殖速率。
1.1 竞争的类型及特点 竞争不外乎包括对生存空间、食物或营养物质、光等资源的争夺。广义的竞争包括捕食、取食、寄生等,物种A要对物种B捕食或取食,物种B并非毫无反应,而是采取相适应的措施来逃避捕食或在体内产生化学物质使不利取食。狭义的竞争是专指两生物具有共同的食物、空间等所产生的竞争关系。通常在同一地区,物种越丰富,种间竞争越激烈。同样在同一空间范围内,同一物种的个体越多,种内竞争也越激烈。从物种关系来考虑,竞争则分为种内竞争和种间竞争。种内竞争是指同一物种不同个体之间对资源的竞争,种间竞争是指两个或多个物种之间对资源的竞争。一般而言,种间竞争的物种具有相同资源需求,也即具有部分或全部相同的生态位幅度。
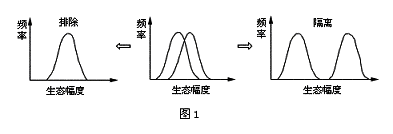
1.2 竞争的结局 在种间竞争中,两个生态上接近的种类的剧烈竞争,从理论上讲可以向两个方向发展(图1),一是一个物种完全排除了另一个物种,使其不能生存。如Gause(1932)所进行的大草履虫Paramecium caudatum和双小核草履虫P.aurelia培养实验;另一是生态隔离,一个物种使另一个物种占有不同的空间(如异域分布)、取食不同食物(如食性上特化)或其他生态习性上的分隔(如活动时间的不同)。如Crombie(1947)所进行的锯谷盗Oryzaephilus surinamensis和拟谷盗Tribolium comfusum实验,这两种昆虫共同生活在面粉中时,由于拟谷盗积极杀死未成熟的锯谷盗而消灭它,但在面粉内放上管子,体形较小的锯谷盗则能逃脱拟谷盗的攻击,两个物种就能共存。

同一物种内,其不同个体的生态要求是一致的,作为一种密度制约过程,种内竞争的效应经常是可测的,如前述的逻辑斯谛增长曲线。Nicholson(1954)认为种内竞争有两种极端类型的竞争现象(图2),他把这两种类型称之为“争夺性”竞争和“分摊性”竞争。在争夺性竞争(又称为干扰性竞争)中,每一个获胜的个体均能得到其所需要的一切物品,而失败者所得的需求品不能满足其生存或繁殖的需要而最终死亡。如某些独立性土蜂对有限数量的巢穴的竞争,某些独居性寄生昆虫对寄主的竞争属于这一类型。在植物方面,用来解释争夺性竞争的例子是植物对阳光和空间的竞争。假如在一苗圃撒播某一大型乔木的种子,出苗后可以见到许多个体,若不迁移随其自身发展,则随着植物的长大,存活的苗木越来越少。在分摊性竞争(又称利用竞争)中,通常获胜者是不完全的,为竞争者取走的部分资源,不能用于维持该种群,资源在所有竞争着的个体间的分摊是均等的,所以当每个个体所摊得的资源不足以维持生存时,死亡率将立即从0突升到100%。如以1g重的公牛脑匀浆作为食物,在不同密度下饲喂培养羊绿蝇Lucilia cuprina幼虫,结果发现在低密度下,羊绿蝇可以发育为成虫,但当密度为200头或更多时,羊绿蝇因分摊的食物资源不足,均不能正常发育而全部死亡。
1.3 对竞争的评价 竞争在动物群落结构中起关键性作用,地理的、形态的和资源利用的种间格局,虽然不是直接的证据,但对发展这一观点是非常重要的。特点是在脊椎动物方面,竞争可以直接观察,如一个物种对另一个物种的空间替代。许多生态学家认为:群落的总的格局归功于种间竞争。
竞争是生态学中的一个重要的过程,是生物进化的一个选择因子(Keen, 1982)。通过竞争使一些相似的物种具有不同的食物、占有不同的栖境、在不同时间取食、占有不重叠的区域或以其他形式分享限制它们数量的因子(Lack,1977)。在同一昆虫群落内,往往可见到多种草食昆虫同时存在,它们的食性各异,这种食性特化可能是竞争反应影响的。但是,也有人怀疑竞争的普遍重要性,他们不认为所有昆虫格局归功于种间竞争。与脊椎动物群落相比较,由种间竞争而确定的格局,在食叶类昆虫是很少的或微弱的,虽然在食叶类昆虫竞争的某些例子被发现,但它们的频率是如此的低,以至于很难把竞争确认为食叶类昆虫群落结构调节的主要过程(Karban,1986)。假如食叶类昆虫之间不存在竞争,或竞争非常微弱,那么,群落的格局应是一致的,然而,从昆虫群落格局看来,情况并非如此。一般而言:1)大的区域比小的区域具有更多的昆虫种;2)复杂的栖境比简单的栖境具有更多物种;3)持久的栖境比短暂的栖境具有更多的生态位窄的物种;4)在同一个同资源集团(guild )内,存在种内或种间个体大小显著的生态差异;5)在生态时间中,群落具有物种“饱和”;6)在非常密切的近缘种之间,物种竞争最激烈;7)在一定栖境内,物种表明“生态位隔离”(Lawton等,1981)。
竞争排除原理强调的是两个生态学上完全相同的物种不可能同时同地生活在一起,其中一个物种最终必须将另一个物种完全排除,如果两个物种实现了共存,那么在它们之间,必然会存在生态学上的差异。这些差异的存在常常用于解释为物种减少竞争的机制。如植食性昆虫种间竞争,虽然不象脊椎动物那样具有明显的敌对行为等竞争形式,但这并不能说明不存在竞争。认为植食性昆虫之间不存在竞争,可能由于:1)植食性昆虫密度通常低于达到种间竞争的种群密度;2)竞争排斥没有发生是由于物种具有不同的地理中心。另外在自然界不可能找到在生态学上需求完全一样的两个或多个物种,这也是否定该假说的原因之一。
2 适应
地球上现存的生物估计有两百多万种以上,而曾经生活在地球99. 9%的物种现在已经灭绝。从6亿年前至今,最保守的估计已有20亿种生物在地球上出现过(Lewontin,1978),为什么会灭绝?显而易见是它们不能适应变化着的环境。达尔文的自然选择进化理论解释生物界两个不同表现方面:分化和适应。分化说明了为什么地球上物种越来越多,而适应说明了现有物种是长期适应的结果,灭绝了的物种则不适应其所处的环境。
2.1 什么是适应 适应有两方面的含义,一是指生物通过变化而能在某一环境中更好地生活的过程;另一是有利于生物在环境中生存和繁殖后代的任何发育上的、行为上的、解剖上的或生理上的特征。从适应的定义可以看出,物种对环境的适应就是为了更好的生存或保存自己。任何一个物种,从它的形成就一直想扩充自己的分布范围,并尽可能使种群达到最大。然而事实上,物种分布有一定的范围,并且在该范围内有一定的数量,并且常常稳定在一定数量之下。这是由于一方面自然界的环境不都是合适的或无法适应,另一方面在适应的环境内,当种群密度高时,自身对环境因子产生了改变,不利于自身生存,或环境容量有限,这样通过竞争等途径淘汰了一些不适应的个体,而使最具适应能力的个体保存下来。
2.2 适应的形式 生物对外界的适应是多方面的,有形态的、生理的、行为的和生态的适应。就陆生动物而言从水生起源发展到陆地生活,要克服的第一个障碍就是水能否满足生物有机体内一切生命活动的基本需要。以昆虫为例说明,昆虫的绝大部分种类是生活在陆地上,从水生到陆生,昆虫经过长期的适应产生了一系列的形态结构、生理过程来适应陆地生活条件。为了减少水分的丧失,体表具有蜡质层作为保护层、控制气门的开闭时间、后肠水分的再吸收、利用代谢水等。一旦适应陆地生活后,动物则对水分状况各异的环境,在行为上采取不同的适应方式。如选择潮湿的小生境、选择一天中湿度适宜的时间出来活动、干旱季节的迁徙、夏眠与滞育等。如生活在干旱荒漠中的动物,大部分种类是生活在地下的,美国西南部的一种大毒蜥一年中大约有98%的时间是在地洞里生活。
2.3 适应有终点吗 自然界并非是静止的,时时刻刻都在不断地变化,生物为了生存在这个地球上,也时时刻刻在不断改变自己,来适应这个变化的自然界,因此,对适应而言,没有终点。
研究物种生存的核心问题是物种的生态位,生态位是生物生存的整个环境及生活方式的多维描述。按照适应的概念,进化被描述为生物对生态位的适应过程,也即生态位在物种适合它之前就已存在,通过新物种的进化来填充。事实上生物并非是被动地感知环境,它们能创造和锁定它们所居住的环境。同时,按照这个理论,生物若占有了其生态位,那么,生物已是适应了这个环境而不需改变。其实现存物种的环境是经常衰败的或变化的。从这个角度出发,物种在长期的适应过程中,自然选择的本质不是提高物种的生存机会,而是简单地使物种“跟踪”或跟上经常变化的环境。
2.4 环境的剧烈变化,生物能适应吗 随着人类工业化的高速发展,技术的不断更新,在地球上已经没有尚未出现人类足迹的地方,人类的干扰、大气污染形成的酸雨、臭氧层的破坏、生境的破碎及退化、生态系统的失调等一切改变着生物的现存空间,在生物圈中的生物有一部分来不及适应或根本无法适应新的环境,将随着环境的剧烈变化而将永远消失。从地球上物种灭绝的情况来看,生物进化史中曾出现过6次物种大灭绝事件(Sepkoski等,1986),对灭绝的原因提出了许多假说,试图加以解释,虽然这些假说至今没有一个经过了严格的论证,但有一条是肯定的,即物种灭绝是由于物种未能适应当时环境的结果。目前生物多样性受到普遍关注,这是由于人们感到对生物多样性的威胁是前所未有的,特点表现在众多物种在如此短的时间遭受灭绝。如果这一趋势继续下去,那么我们面临的灭绝规模不亚于历史上任何一次物种大灭绝。人们意识到这次灭绝必然会危及人类自身。为了保护生物多样性,大量的物种在人工措施下饲养或栽培,它们在人类的悉心的管理下尚能成长发育,一旦置于野生的自然环境,则根本无法适应或很难适应,变成了脆弱的物种。因此有人预言未来的野生生物将在人类的集约管理下生存 ,这是一个可怕预言。一些种类(如鼠类、蟑螂等)适应了人类改变或创造的环境而大量地繁殖起来形成灾害。
颜忠诚,生物学通报,2000年第3期
扩展资料
植物的防御种种
植物在长期与自然环境的斗争中,形成了保证物种生存的各种各样的防御办法。
一、利用毒素防御
植物能产生多种有毒化合物,如生物碱、葡萄糖甙、树脂类物质和有机酸等。这些毒素是植物最有效的防御武器。当植物在被触摸或吃掉时,这些毒素便发挥有效的作用。如常春藤分泌生物碱,使人和动物起皮疹。高梁、箭草和某些野生植物分泌氢氰酸使人和动物在大量吞食后中毒。还有些植物如荞麦、金丝桃等分泌出“光敏毒素”这种有害物质,它使人和动物对光极度敏感。一旦动物误食含有这种毒素的植物后,在遮阴处没有什么反应,但在强烈的阳光下,立即发生多种症状,甚至造成死亡。有些植物如毒草、野紫菀等本身不制造毒素,而是直接从土壤中吸收硒等有毒元素,使动物引起“急性硒中毒”。有趣的是,含有毒素的植物并不是各个部分都有毒,大部分毒素集中在植物最易受袭击的部位,如叶、花、果。
二、利用分泌物防御
某些植物如报春花、虎耳草等从组织中分泌出很粘的分泌物散布在植物的表面,以捕捉那些爬上来的昆虫,这是一种十分普遍的有效防御。这种分泌物可能产生于表面成特殊的分泌腺和毛茸。这种分泌物对本身的保护不仅仅是花,而且也扩展到整个簇叶上。还有植物如凤生花属植物,从叶基部托叶上发育出的特殊分泌腺分泌出一种能吸引昆虫爬上茎的“甜流”,昆虫遇到这种甜物质就停止前进,从而使花蜜安然无恙。
三、利用难闻的气味和味道防御
某些植物如木贼的茎、杜鹃花的叶等积存了大量的二氧化硅,使动物吃后不易消化。还有些植物如胡椒、辣椒的叶子等,含有多种不可口或刺激性物质,或酸或苦,大多数动物尝过以后就不再问津了。由于多数草食动物不喜欢叶面上有水珠和露水的植物,有些植物发展了一种保持由露水形成的水珠时间比其他植物更长的能力。如川续断科和菊科植物,从而达到保护自身的目的。
四、利用针、刺和荆棘等特殊结构防御
某些植物如仙人掌、小蘖等,叶变成刺,皂类、枳等枝变成刺,一方面植物利用这些刺保护自己,另一方面可以减少水分的蒸发。
某些植物如荨麻把针和毒这两种防御武器相结合,从而产生更有效的保护作用。它的倒刺几乎起皮下注射器的作用,即能戳入侵害它们的动物的皮肤,同时毛刺从叶子上脱落下来,牢牢地扎在皮肤上,并把使人不舒服的毒素注入皮肤中,引起皮肤严重的炎症。
针、刺、荆棘像植物的毒素一样,都是策略性地分布在植物体上,以叶、花、果上最多。它们排列在动物取食时首当其冲的位置上。这些保护结构的分布与大小,取决于最可能侵害植物的动物种类及最易受害的植物部位。如浮在水面上的王莲的大叶子仅仅在下表面和边上有保护结构,因为这些部位最易受害。松树的针叶,芦荟的针状、窄长、坚硬和带尖的叶子、刺草的锯齿状叶缘,能刺伤动物的嘴和内脏器官,所以动物并不伤害它们。
五、利用巨大的数量防御
有些植物如禾本科植物完全没有防御能力,但由于它们生长快、繁殖茂盛,才很少遭受牛、羊、马等草食动物彻底毁灭性的危害。
有些植物还利用拟态、共生等方式保护自己。
陈瑜,中学生物学,1998年第3期
扩展资料
浅谈适应
适应是生物适合环境条件而形成的一定特征和性状的现象。它包括两重意义:一是指过程,即生物不断改变自己,使其能适合于某一环境中生活的过程。例如肺鱼的鳔在缺少水的时候进行呼吸,摄取氧气以维持生命。二是指结果,即有利于生物生存和繁殖的种种特征。例如植物的叶适应于光合作用;鱼的鳃适应于水中营呼吸作用。
适应的现代观念认为:外部世界提出某些“问题”,生物就必须“去解决”,生物通过对“问题”的解决过程作出越来越好的“回答”,最后处于适应状态。爬行类演化成鸟类就是一个很好的例证。在演化的过程中,骨胳、肌肉、皮肤、翅膀、羽毛等越来越变得更适应于鸟类的生活。
生物适应于自然环境,具有普遍性。生物只有适应于其所处的环境,才能够在这个环境中很好地生存下来。各种植物花的形态、构造都有利于其后代的繁衍。虫媒花的构造、颜色、花蜜、香气等特征极易吸引昆虫为之传粉受精、风媒花花粉粒小而轻且数量大,这些特征与风的传粉相协调;蒲公英的种子上生有毛绒绒的白色纤维,适于被风吹走;苍耳和鬼针草等植物的种子依靠动物来传播,种子、果实上有刺,很容易附着在动物身上;风仙花种子成熟后,被风一吹,立即裂开,里面的种子便被弹射出来。
1844年,邦尼艾(G・Bonnier)做了一个实验,将生长在同一个地方的菊芋一部分放在平原上生长,一部分放在海拔2400米的植物园里,经过数代后,在植物园里的菊芋发生了巨大的变化,其形态、结构、生理机能变得更适应于高原生活了。
在动物方面,保护色、警戒色及拟态等现象都是动物对其周围环境的适应。枭蝴蝶身上斑点的颜色和形状与猫头鹰的眼睛十分相似,当它突然展开翅下的斑点时,可即刻恐吓并赶走捕食者;竹节虫,整天不动,犹如枯枝败叶,如震动枝条,竹节虫便跌落在地,仍然僵直不动,活象一段枯枝;蛇、鸟等动物在地震前都自行逃走;蝙蝠的翼手适于飞翔;鲸、海豹、企鹅的某些器官适于划水等等。
总之,适应是普遍存在的生命现象,各种生物,无论是形态、结构,生理机能以及行为习惯,无一例外。
适应是自然选择的结果,生物的变异有的有利于生存,有的不利于生物的生存。那些有利于生物生存的变异类型适应自然环境而留下来;那些不利于生物生存的变异类型由于不适应自然环境而遭淘汰。所以,适应是自然选择的必然产物。
适应现象尽管很巧妙,很合理,但适应是相对的,而不是绝对的、尽善尽美的。适应是针对一定的条件说的,如果条件发生了改变,那么,生物的适应就失去了作用。警戒色是相当巧妙的适应现象,大多数食虫鸟怕吃毛虫,但是一只杜鹃在一天中却可以吃掉上百条毛虫。蛾类结茧固然有利于保护自身,然而棉铃虫的悲剧又恰恰就在茧上,金小蜂正是借助于茧对棉铃虫的束缚作用,顺利地将卵产到它的体内这也可以说是“作茧自缚”了;鹰可以在很高的地方发现草丛中活动的老鼠,人的眼睛则不行。相反,人的眼睛可以辨别多种颜色,鸟类的眼睛则不行。
适应是一种暂时的现象,它不是永久性的,当某些因素变化时,有的还成为一种有害的特征,导致生物死亡。雷鸟和貂都借助于保护色而常常免遭敌害。但是一旦这些动物的颜色已经变了,而气候异常没有下雪,它们的颜色变更,不仅没有益处,反而更容易被敌害所发现。著名的尺蛾工业黑化现象也是一个很好的例子。
适应相对性究其原因是由于遗传基础例如基因等的稳定性和自然环境条件的变化相互作用的结果。环境发生了剧烈变化,适应于原来环境的生物绝大多数要被淘汰,但其中的一些生物由于遗传物质发生了变异,能够在新的环境中生活下来,适应于变化的环境。例如在一个生长着细菌的培养器中,加入青霉素,绝大多数细菌不适应而被杀死,但总有一些发生变异的个体,抗药性增强而适应于新的环境生存下来。
李锦桂,中学生物学,1991年第4期